γδT细胞与牛皮藓(银屑病)
一.γδT简介
γδ T细胞是表面具有独特T细胞受体(TCR)的T细胞。大多数T细胞是αβ T细胞,其TCR由两个糖蛋白链组成,称为α和β TCR链。相比之下,γδ T细胞具有由一个γ 链和一个δ 链组成的TCR。αβT和γδT细胞谱系起源于缺乏CD4和CD8共受体(CD4-CD8-)的T前体细胞,也称为双阴性(DN)胸腺细胞(图1)。γδT细胞通常不如αβ T细胞常见,但在粘膜和上皮部位显着富集,例如皮肤和呼吸道,消化道和生殖道。与αβT细胞相比,γδT细胞是主要的组织相容性复合物(MHC)非限制的先天样淋巴细胞,具有更独特的抗原受体,产生细胞因子如IL-17 / IFN-γ / IL-22。同时γδT细胞桥接了先天性和适应性免疫系统。虽然它们只占总T细胞的一小部分,但在修复宿主组织,病原体清除,肿瘤监测和炎症调节等方面具有多种作用。

图 1 .γδT细胞的胸腺选择和发育
- γδT细胞分布及作用
人类γδT细胞可以根据δ链表达来区分,其中包括Vδ1,Vδ2和Vδ3亚型。
Vδ1 T细胞主要存在于肠道上皮、皮肤、脾脏和肝脏中,参与维持上皮组织的完整性。它们约占外周血(PB)中γδT细胞的30%。Vδ1T细胞在维持屏障组织完整性和建立抗病毒免疫中起着至关重要的作用。研究表明,Vδ1细胞参与几种疾病,如疟疾,人类免疫缺陷病毒(HIV),巨细胞病毒(CMV),炎症性肠病和克罗恩病通过其细胞毒性和分泌细胞因子发挥作用。值得注意的是,活化的Vδ1T细胞通过NKp30识别B7-H6.B7-H6是仅在肿瘤细胞上表达的B7家族成员,并参与抗肿瘤作用。
Vδ2 T细胞主要分布在血液和淋巴系统中,是健康人类中发现的主要亚群。它占外周血γδT细胞群的50%-90%。Vδ2T细胞分为先天样(Vγ9 +)和适应性(Vγ9-)亚群,大多数Vδ2T细胞为Vδ2Vγ9 + T细胞。Vδ2Vγ9 + T细胞对细胞因子有反应,例如CCR1,CCR2,CCR5和CXCR6配体以及IL-12,并产生促炎因子,例如IFN-γ,TNF-α,IL-17,IL-21和IL-24。此外,活化的Vδ2Vγ9+T细胞获得抗原呈递细胞(APC)特性,并表现出很强的细胞因子分泌能力,如Th1/Th2 /Th17型细胞因子。这些诱导树突状细胞(DC)成熟为APC。
Vδ3T细胞是外周血淋巴细胞的最小亚群,占循环细胞的0.2%。它们表达CD56,NKG2D,CD28,HLA-DR,CD161和T细胞活化标志物CD69,但不表达CD25,NKG2A或NKG2C。Vδ3T细胞在肝脏和肠道中是丰富的,并且参与慢性病毒感染和白血病。扩增的Vδ3T细胞仅识别CD1d并释放Th1,Th2和Th17细胞因子以诱导树突状细胞成熟成APC。它们不识别 CD1a、CD1b 或 CD1c 。Vδ3 T细胞和B细胞相互调节成熟标志物CD40,CD86和HLA-DR的表达,并促进B细胞的IgM释放。
此外,在淋巴瘤患者的PB中观察到Vδ4,Vδ6,Vδ7和Vδ8 T细胞,但它们的作用机制尚有待阐明。
三.γδ T细胞在免疫介导的皮肤病(银屑病)中的作用
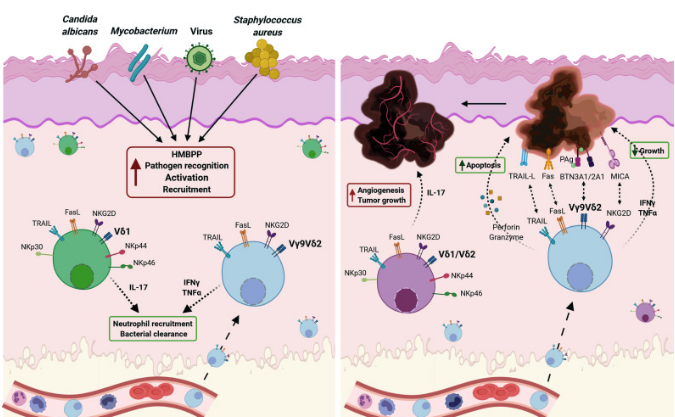
循环Vγ9Vδ2 T细胞在感染多种病原体后可立即招募到皮肤上,包括病毒,分枝杆菌,白色念珠菌和金黄色葡萄球菌(图2左)。病原体活化的γδ T细胞促进细菌清除和嗜中性粒细胞募集,从而参与疾病的解决。Vδ1 T细胞的数量在白色念珠菌感染中扩增,它们分泌IL-17。在癌症中(图2右),Vγ9Vδ2 T细胞中表达的不同受体对恶性抗原的直接识别可以诱导IFN-γ和TNF-α的分泌,也可以介导肿瘤细胞的直接裂解。因此,Vγ9Vδ2 T细胞是控制肿瘤生长和扩张的有效工具。肿瘤微环境中γδ17 T细胞的存在有助于肿瘤血管生
成和生长,促进癌症的进展,是免疫治疗的重要靶点。
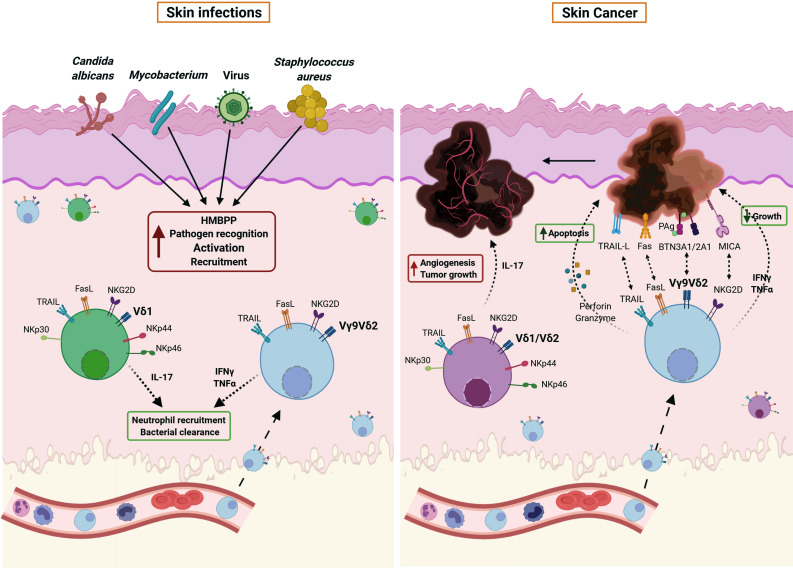
图 2 .γδT细胞在皮肤感染和肿瘤中的作用
免疫系统失调和T细胞活化已被充分证明在银屑病发展中起着至关重要的作用。一些研究将皮肤中的T细胞功能归因于αβT细胞,而γδT细胞经常被忽视。IFN-γ产生T辅助(Th)1细胞最初被认为是银屑病的主要驱动因素。然而,过去十年的大量临床和基础研究结果已经证明白细胞介素(IL)-23 / Th17轴在银屑病的发病机制中起重要作用。发现在IL-23-和IL-17缺陷小鼠银屑病炎症出现进展导致损伤,从而证实了IL-23 / IL-17轴的参与。Th17细胞及其下游效应分子,包括IL-17A,IL-17F,IL-22和肿瘤坏死因子(TNF-α),在血清和银屑病皮肤病变中增加。最近,Th17细胞被发现不是银屑病中这些致病细胞因子的主要来源。相反,发现IL-17A,IL-17F和IL-22是由γδT细胞产生的。将IL-23注射到小鼠的皮肤中或应用局部剂量的咪喹莫特乳膏(5%)诱导典型的银屑病样表型,即表皮厚度,红斑和炎症。这两种模型被证明可以模仿银屑病样炎症,并已被用于评估不同治疗方法的疗效。用这些模型研究数据表明,真皮γδT细胞是皮肤中对IL-23刺激产生IL-17的主要细胞。
真皮γδ T细胞产生IL-17需要内源性IL-1β。在机理上,IL-1β通过IL-1R-MyD88激活雷帕霉素(mTOR)信号通路的哺乳动物靶标,而IL-23激活STAT3通路。转录因子IRF-4将IL-1R和IL-23R途径联系起来,以诱导真皮γδ T细胞中增强的IL-17产生。Vγ4和Vγ6真皮T细胞都产生IL-17,然而,与Vγ6相比,真皮Vγ4 T细胞扩增并产生显着更多的IL-17。真皮Vγ4和Vγ6T细胞具有不同的效应子信号传导要求。真皮Vγ4 T细胞增殖和IL-17的产生依赖于STAT3,而真皮Vγ6 T细胞可能通过STAT3非依赖性RelA / NF-kB途径被激活。因此,真皮Vγ4 T细胞似乎在IMQ诱导的银屑病样皮炎中起关键作用。
真皮γδT细胞表达IL-23R,IL-17R,RORγt和趋化因子受体CCR1,CCR2,CCR4,CCR5,CCR6,CXCR3和CXCR4。CCL20是一种独特的CCR6配体,介导产生IL-17的γδT细胞和DC的皮肤浸润。大量研究表明,CCL20 / CCR6调节γδT细胞从真皮到表皮的迁移,促进中性粒细胞聚集,并加剧炎症。在IL-23注射的WT小鼠中,CCL20被高度上调,在表皮中观察到许多CCR6 + γδT细胞。具有最小趋化活性的抗CCL20中和抗体或工程CCL20适配体可防止产生IL-17的γδT细胞浸润到IL-23注射小鼠的皮肤中。这导致IL-17和IL-22下调,阻断γδT细胞募集到表皮,并减少银屑病样皮炎。在CCR6-敲除(KO)小鼠中,γδT细胞在IL-23处理后未能迁移并积聚在表皮中。这表明CCR6 / CCL20作为银屑病治疗靶点的潜在相关性。
银屑病频繁发作,无法根治,治疗停止后在同一区域发生复发。因此,复发性银屑病是一个需要解决的重大问题。TNF-a,IL-12 / 23和IL-17抑制剂已被证明表现出有效和快速的治疗效果。然而,这些生物制剂与几种不良事件有关,最常见的是对感染的易感性。除感染外,生物抑制剂还与脱髓鞘疾病、鼻咽炎、上呼吸道感染、头痛、狼疮或狼疮样综合征、皮肤黏膜念珠菌病、轻度中性粒细胞减少症以及心力衰竭新发或恶化有关。长期的安全问题和高成本阻碍了这些试剂的广泛使用。
原始病变区域周围的银屑病复发表明这些表现具有“免疫记忆”。在银屑病小鼠模型中已经观察到记忆样γδT,产生IL-17A的Vγ2Vδ4 + T细胞最初来自新生儿胸腺,在那里它们被指示具有组织向性。这些Vγ2Vδ4 +T细胞具有表型记忆样,具有CD44hi CD62Llo CD27-表达模式。暴露于IMQ后,皮肤中的Vγ4 + γδT17细胞已被证明在引流淋巴结(LN)中迅速扩张,然后从LN中释放出来。然后,它们通过趋化因子CCR2的作用迁移,以S1P1依赖性的方式积聚在发炎和未发炎的皮肤部位。这反过来又加剧了中性粒细胞的炎症反应和募集。它们也被证明可以通过血液迁移,并在正常皮肤和外周LN中持续至少三个月。重要的是,当在远处的皮肤部位接受相同的第二次刺激时,与暴露于第一次刺激后相比,记忆样Vγ4 + γδT17细胞以更快的速度膨胀并产生更多的IL-17,导致快速和严重的皮肤炎症反应(图3)。致敏小鼠在IMQ激发时表现出皮肤炎症升高,细胞增殖显着,Vγ4 + γδT细胞产生IL-17。过继转移实验已经证实,记忆样Vγ4 + γδT17细胞反应迅速,并且它们的记忆驱动它们参与银屑病复发。
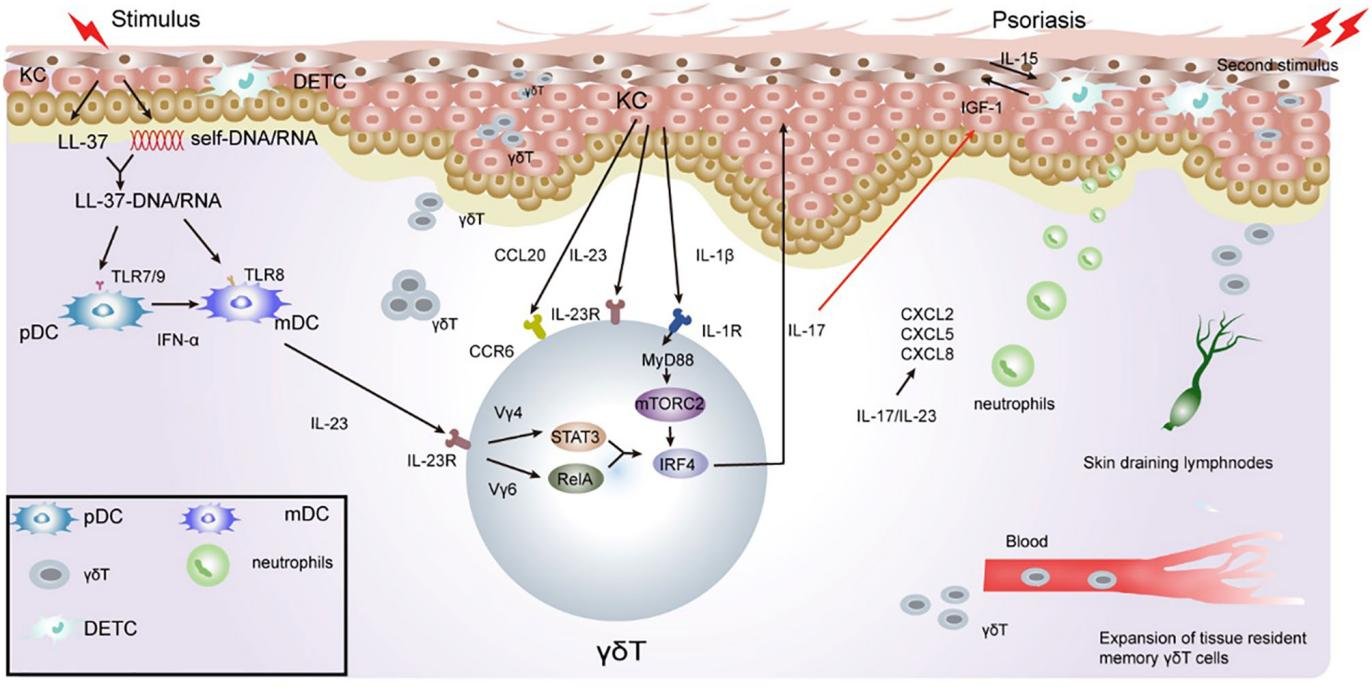
图3γδT细胞在银屑病免疫发病机制中的作用以及记忆性
四.靶向γδT细胞进行银屑病治疗
真皮γδT细胞在银屑病发病机制中的重要作用在过去几年中已经阐明。因此,真皮γδT细胞及其相关分子已成为药物开发的有吸引力的靶标。脂联素是胰岛素敏感性的代谢介质,在代谢调节和炎症/抗炎过程中起着至关重要的作用。研究表明,在没有脂联素的情况下,在银屑病样皮肤中,产生IL-17的真皮γδT细胞的炎症和浸润显着增强。脂联素对真皮γδT细胞产生IL-17的负调节主要通过AdipoR1介导。这表明增加脂联素水平可能对改善银屑病以及代谢紊乱有效。BTLA属于免疫球蛋白超家族,据报道在淋巴组织中γδT细胞/ ILC的稳态中起作用,并控制成熟淋巴结γδT细胞中IL-17的产生。BTLA缺乏的动物模型已被证明具有比例失调的炎症γδT细胞,并且容易患上银屑病和严重的皮肤炎症。发现BTLA激动剂限制了这些表型的进展。BTLA的激活可以恢复γδT细胞亚群的平衡以控制自身免疫发病机制。抗激动性抗BTLA抗体(clone 6A6)被证明抑制IMQ诱导的淋巴结和皮肤内的γδT细胞扩增和IL-17的产生。因此,BTLA可能是治疗银屑病的潜在靶点。真皮γδT细胞表达CCR6。在IL-23诱导的皮肤炎症小鼠模型中,CCR6基因敲除或使用抗CCL20单克隆抗体导致银屑病性皮炎的下降。这表明CCL20及其受体CCR6是治疗银屑病的潜在靶标。CCL20 S64C是CCL20变体,与CCR6结合并抑制CCR6介导的γδT细胞迁移。先前的研究表明,CCL20 S64C减轻了IL-23诱导的银屑病样模型中的炎症反应,并且与CCR6 + IL-17产生γδT细胞在表皮中的积累减少有关。FTY720是FDA批准的用于治疗多发性硬化症的免疫调节药物。它通过抑制鞘氨醇-1磷酸受体(S1PR)来减少淋巴细胞从淋巴组织的排出。FTY720抑制Vγ4 + VγT4 + T17细胞从淋巴结到皮肤的迁移,表明其作为银屑病治疗的潜力。靛红素(IR)是从中国草本植物Indigo naturalis的叶子中提取的双吲哚化合物。已经证明,通过主要通过减少IL-17 A 生成 γδT细胞来缓解IMQ诱导的银屑病样皮炎。Dashlkhumbe等人报道了一种新配制的甲氨蝶呤(MTX,MTX与细胞渗透性肽的化学偶联物)用于治疗银屑病。局部应用皮肤渗透(SP)-MTX通过减少CD11c +,CD4 +和IL-17产生γδT细胞的皮肤免疫细胞浸润来减少银屑病样皮肤现象和表皮厚度。
五.总结
银屑病具有复杂多样的发病机制。在疾病发展过程中,γδT细胞分泌促炎细胞因子,如IL-17和IFN-γ,诱导和加重银屑病。值得注意的是,γδT细胞具有对二次刺激快速反应的记忆细胞特性。这会促进银屑病的复发。
未来的研究应该调查存在于皮肤病变中的γδT细胞是否具有常驻记忆细胞特性,它们持续多长时间,它们翻转的频率,以及外周组织中的哪些环境位支持它们的长期生存。需要进一步的研究来确定减少γδT细胞以防止银屑病复发的机制。
咨询
- 357
- 点赞
- 复制链接
- 举报