

用于长时间药物检测的神经球和肝脏类器官培养物的多器官串联共培养
目前用于药物开发的体外和动物试验未能模拟人体全身器官的复杂性,因此往往不能准确预测药物毒性,导致临床研究中的高损耗率。人类和实验室动物之间的系统发育距离是巨大的,这影响了神经保护药物疗效的动物数据的可转移性。因此,许多在动物身上显示出希望的神经保护治疗在转移到人类身上时并不成功。我们提出了一种多器官芯片,能够在组合介质电路中维持来自不同细胞来源的3D组织,这弥补了系统和人体测试的空白。在多器官芯片
用于长时间药物检测的神经球和肝脏类器官培养物的多器官串联共培养
A multi-organ chip co-culture of neurospheres and liver equivalents for long-term substance testing
Eva-Maria Materne a,∗,1, Anja Patricia Ramme a,1, Ana Paula Terrasso b,c, Margarida Serra b,c, Paula Marques Alves b,c, Catarina Brito b,c, Dmitry A. Sakharov d, Alexander G. Tonevitsky e, Roland Lauster a, Uwe Marx a,f
a Technische Universität Berlin, Institute of Biotechnology, Gustav-Meyer-Allee 25, Berlin 13355, Germany
b iBET, Instituto de Biologia Experimental e Tecnológica, Oeiras 2780-901, Portugal
c ITQB-UNL, Instituto de Tecnologia Química e Biológica, Universidade Nova de Lisboa, 2780-157 Oeiras, Portugal
d Scientific Research Centre Bioclinicum, Ugreshskaya ul. 2/85, 115088 Moscow, Russia
e Institute of General Pathology and Pathophysiology, Russian Academy of Medical Science, Baltiyskaya ul. 8, 125315 Moscow, Russia
f TissUse GmbH, Markgrafenstr. 18, 15528 Spreenhagen, Germany
翻译整理:北京佰司特贸易有限责任公司
背景介绍
目前用于药物开发的体外和动物试验未能模拟人体全身器官的复杂性,因此往往不能准确预测药物毒性,导致临床研究中的高损耗率。人类和实验室动物之间的系统发育距离是巨大的,这影响了神经保护药物疗效的动物数据的可转移性。因此,许多在动物身上显示出希望的神经保护治疗在转移到人类身上时并不成功。我们提出了一种多器官芯片,能够在组合介质电路中维持来自不同细胞来源的3D组织,这弥补了系统和人体测试的空白。在多器官芯片中,将人人工肝类器官和人神经球置于流体中两周的稳态类器官串联共培养成功地证明了其长期性能。每天测量乳酸脱氢酶活性和免疫荧光终点染色证实了组织的活力和分化细胞表型的持续维持。此外,培养组织的乳酸产量和葡萄糖消耗值表明,在类器官串联共培养6天后,达到了一个稳定的稳态。通过qPCR和神经元标记物βIII -微管蛋白和微管相关蛋白-2的免疫荧光证实,在多器官芯片中培养两周后,神经球仍保持分化神经元。此外,在两周的毒性试验中,重复暴露于两种不同浓度的神经毒性2,5-己二酮物质,可诱导神经球和肝脏类器官内的高凋亡,培养液中乳酸脱氢酶活性显著增加。对2,5-己二酮类器官串联共培养的主要发现是,不仅可以区分两种不同剂量的毒性谱,而且与多器官芯片中各自的单类器官培养相比,类器官串联共培养对该物质更敏感。因此,我们在这里提供了一种新的体外工具,可以在未来的临床研究中更准确地预测药物的安全性和有效性。
研究介绍
新药的开发非常昂贵、耗时,而且只有少数几种药物能经受得住漫长的试验阶段。动物实验往往无法预测新药的效果,因为实验动物和人类之间的系统发育距离太大(Bailey et al., 2013;Dragunow, 2008)。大量的实验动物用于物质测试不仅昂贵和费力,而且在伦理上也有问题。因此,必须找到新的技术来在过程的早期阶段评估潜在的候选药物。微流控培养装置结合人体的类器官在一个类似器官的安排,在稳态稳态下很可能成为一个翻译解决这一测试困境。 目前,能够在体内模拟人体情况的微流体平台非常少(Hwan,2009;2009)。这些系统的缺点大多是外部泵和外部介质容器,导致较高的液-组织比。因此,不能保证组织之间的交流。此外,大多数微流控系统中的物质或药物的暴露时间在24 - 72小时之间,只有少数可以长达7天(Marx et al., 2012)。然而,经济合作与发展组织(Organization for Economic Co-operation and Development,简称oecd)在动物身上对化学品和化妆品进行毒性测试的指导方针要求28天的重复剂量的暴露。
在此,我们评估了我们实验室开发的微流控多器官芯片(MOC)平台是否能够长时间类器官串联共培养神经球和肝脏类器官。MOC覆盖了物体载玻片的区域,包含一个芯片上的微泵,并能够互连两个不同的类器官培养物。该微型泵可确保介质以可变流速通过组织培养室的稳定长期循环,可调节以模拟各自组织中依赖血流的机械剪切应力(Schimek,2013)。培养室和连接通道是光学可达的,因此,支持活组织成像。我们在这里展示了MOC能够支持人肝脏类器官的类器官串联共培养,包括分化的肝parg细胞和人肝星状细胞,以及分化的神经球,来自NT2细胞系,在一个联合的培养基回路中,在数周的稳定状态下。通过免疫荧光染色和所选标记基因的qRT-PCR检测,细胞的分化表型得以保留。此外,系统布局和芯片设计支持为安全性或有效性测试试验开发而重复的药物的接触。
在为期两周的毒性试验中,2,5-己二酮被用来研究它对组织的毒性作用。在人的肝脏中,正己烷要么通过解毒途径代谢为1-己醇或3-己醇,要么通过生物活化途径代谢为2-己醇。2-己醇在许多氧化步骤中进一步代谢为2,5-己二酮和其他代谢物(Yin et al., 2013)。这些物质被分配到血液中,到达肾脏和大脑等其他器官。2,5-己二酮是导致神经毒性的主要成分,因为与其他代谢物相比,它在人体内的滞留时间最长。2,5-己二酮可以通过吡咯加合物的形成和共价交联来改变神经纤维的结构(Heijink et al., 2000)。对实验室动物和细胞系的不同研究表明,神经毒性作用(DeCaprio等,1988;Ladefoged,1994)。然而,2,5-己二酮对神经元细胞(NT-2- n, SK-N-H)和非神经元细胞(NT-2) 4小时的暴露没有显示任何细胞毒性,但32 mM 2,5-己二酮24小时的暴露导致对两种细胞类型的毒性作用(Woehrling,2006年)。
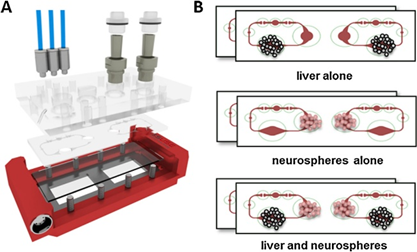
图1:多器官芯片。(A)装置的示意图,包括装有两个微流控电路的PDMS玻璃芯片。(B)神经球和肝脏类器官在MOC中单独培养和类器官串联共培养的实验装置(n = 4)。
表征神经球
神经球从NT2未分化细胞中以三维聚集体的形式在搅拌悬浮培养体系中可重复分化,形成神经球培养典型的致密球形结构(Serra et al., 2009),中等直径为179.0±76.5μm。在培养过程中,球形体的结构完整性保持不变,细胞活力高,无坏死核的迹象,这从广义FDA染色中可以看出,球形体中分布着少量pi阳性细胞(图2b)。
在分化过程中监测培养以表征nt2来源的细胞。nestin(未分化的NT2细胞和早期神经上皮祖细胞的标志)的表达在第10天上调了4倍(图2f)。同时,神经元标记物βIII -微管蛋白的表达增加了8倍。 在第3 - 10天观察到(图2g),可能对应于nestin和βIII -微管蛋白阳性神经元祖细胞的增殖。Nestin的表达从第10天开始下降(图2f),而βIII -微管蛋白保持高表达水平(图2g),表明其向神经元谱系的高效分化。第24天的免疫染色显示βIII -微管蛋白阳性细胞的广泛网络(图2c)。此外,在整个神经球中检测到map2阳性细胞,表明这些神经元正在获得成熟的神经元表型(图2d)。突触前囊泡糖蛋白synaptophysin的基因表达在分化的24天内增加了6倍(图2h);此外,该蛋白在神经礼中被检测到,呈点状模式,与突触前囊泡富集一致(图2e)。map2阳性神经元的检测和突触素基因在分化过程中表达的增加,以及突触素阳性囊泡的积累,提示神经元分化发生在神经球内。
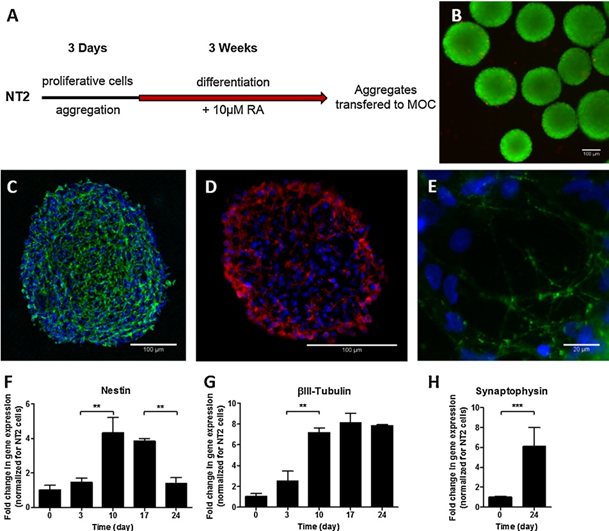
图2:神经球培养的制备、分化和表征。(A)搅拌悬浮培养流程示意图。将细胞接种于搅拌的悬浮培养体系中,聚集三天。维甲酸诱导小鼠神经分化3周。(B)代表ra诱导分化结束时培养状态的图像:荧光活/死试验,使用二醋酸荧光素识别活细胞(绿色)和碘化丙啶识别死细胞(红色)。(C - E) ra诱导分化结束时神经球免疫染色:(C)βIII -微管蛋白(绿色),(D) MAP2(红色)和(E) synaptophysin(绿色)。细胞核用DAPI染色(蓝色)。(F-H) qRT-PCR分析(F) nestin, (G)βIII-tubulin和(H) synaptophysin基因的表达。与NT2未分化细胞相比,神经球mRNA浓度在培养过程中发生了成倍的变化;数据为三个独立培养的均值±标准差。Mann-Whitney检验,星号表示差异显著(P < 0.01)。比例尺B、C、D: 100μm, E: 20μm。
由HepaRG和HHSteC产生的肝脏的特征
如Gripon(2002)所述,在单层培养中预先分化HepaRG细胞。这些细胞以其在培养过程中形成胆管结构的能力而闻名,它们将羧基二醋酸荧光素排泄到细胞间明确的空间中,提示有功能的胆管(图3b)。细胞分化后,与HHSteC联合产生肝脏类器官。
悬滴类器官串联共培养2天,可反复形成中等直径300-400μm,高度200-300μm的肝脏类器官。对新形成的细胞角蛋白8/18和波形蛋白进行免疫荧光染色,发现hhstec均匀分布于整个球体(图3c)。MRP-2在肝细胞中被选择性地转运到胆管的顶膜。在这里,MRP-2在细胞周围的存在表明,生成的类器官中的细胞已经开始像之前在单层培养中观察到的那样改变小管状结构。然而,染色较弱,提示胆管重构不完全(图3d)。此外,功能标记蛋白白蛋白、小管运输蛋白胆盐出口泵(BSEP)、尿素循环酶氨甲酰磷酸合成酶1 (CPS-1)和I期代谢细胞色素P450 (Cyp)酶的qPCR分析显示,向肝脏谱系分化后,其数量增加了20倍。保持在球体中(图3e-k)。与单层分化培养相比,只有Cyp 3A4在球形细胞中的表达显著降低。这可能是由于球体不再用DMSO处理,一种已知的Cyp 3A4诱导剂。然而,利福平3天的治疗显示Cyp 3A4和2B6酶在球状体中的表达是可诱导的(图3i,k)。
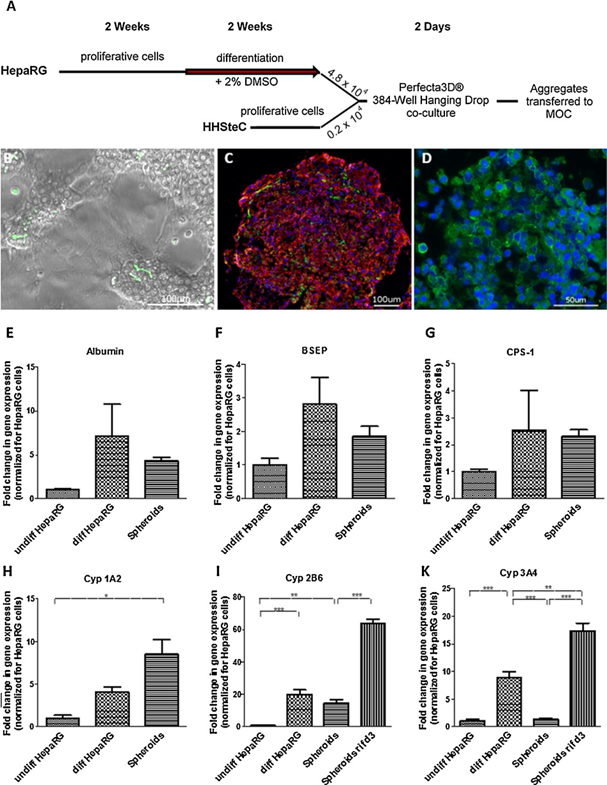
图3:肝脏类器官的制备、鉴别和鉴定。(A)肝脏类器官体制作流程示意图。将HepaRG细胞分化,与HHSteC混合,聚集2 d。用二甲基亚砜(DMSO)诱导肝细胞分化2周。(B)经二醋酸羧荧光素处理的单层肝素显微镜图像。绿色荧光显示胆管。C, D聚集后代表性聚集物的免疫染色:(C)波形蛋白(红色)和细胞角蛋白8/18(绿色)和(D) MRP-2(绿色)。细胞核用DAPI染色(蓝色)。E.白蛋白、F. BSEP、G. CPS-1、H.细胞色素P450 1A2、I.细胞色素P450 2B6和K.细胞色素P450 3A4基因表达的qRT-PCR分析与未分化的HepaRG细胞(undiff HepaRG)相比,分化的HepaRG细胞(diff HepaRG)、新形成的球形细胞(Spheroid rif d3)和球形细胞(Spheroid rif d3)的mRNA浓度发生了显著变化;数据为四个独立培养的均值±标准差。星号表示差异显著(*p≤0.05, **p≤0.01, ***p≤0.001),采用Tukey 's post多重比较检验进行单因素方差分析。比例尺B、C: 100μm, D: 50μm。
MOC中神经球-肝脏类器官串联共培养超过14天
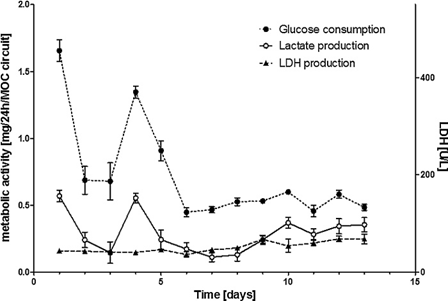
在MOC系统中成功地类器官串联共培养了14天的神经球和肝脏微炎。在整个培养14天的过程中,培养基上清液中恒定的LDH活性表明了一个稳定的组织周转和细胞活力(图6)。葡萄糖消耗和乳酸生成被测量为MOC代谢活动的指标。在类器官串联共培养的第1天,两者均呈下降趋势,在第6天之后达到稳定状态,波动幅度较小。在灌注的MOC中,神经球在培养过程中改变了形态,附着在芯片的玻璃底部。神经球生长,融合在一起,并在两周内形成了一个几乎多层的结构。1.5 Hz的剪应力促进了球的均匀化和融合。肝脏类器官在两周的培养过程中,形态变化不大,保持了3D的球形结构,但也与玻璃底部连接,并与其他球形部分融合。 神经球标本的终点法免疫组织染色,神经元标记物MAP2和βIII -微管蛋白高度阳性(图4d,e)。染色强度与未在MOC中培养的对照组神经球在第0天相当。胚胎干细胞标记物TRA-1-60在所有样本中呈阴性(图4f)。肝脏类器官状细胞角蛋白8/18和波形蛋白染色显示HHSteC仍然均匀分布在整个球状体中(图4a)。此外,细胞色素P450 3A4和MRP-2染色强烈,表明其表型分化良好(图4b,c)。qRT-PCR分析支持这些发现,并显示功能标记物白蛋白、细胞色素P450、2B6和3A4的表达在MOC中培养14天时增加了5 - 15倍(图5a-c)。与神经球类器官串联共培养对肝脏的分化状态没有任何显著影响(图5a-c)。此外,与第0天的球相比,MOC培养中神经球的分化状态保持不变,如nestin,βIII-tubulin和10月4日的表达(图5d-f)所示。
MOC中神经球-肝脏类器官串联共培养的毒性研究
用2,5-己二酮处理肝脏培养物和神经球MOC类器官串联共培养物,观察毒性损伤期间的组织相互作用。对照组MOCs未加该物质处理,在整个培养时间中,培养基中LDH活性恒定在50 U/L左右(图7a)。这些值与处理过的培养物的值一致,直到第6天,当每天开始添加物质,再次表明一个稳定和可行的多组织类器官串联共培养。2,5-己二酮对神经球和肝脏物类器官串联共培养的剂量依赖性效应,在分别用32mm或16mm处理的培养物中,从第7天或第9天开始,LDH活性显著增加。第10天,32m2,5 -己二酮处理的LDH浓度再次下降,表明大部分细胞已经裂解。16 mM处理后,LDH活性的增加延迟了2天,但在达到最高200 U/l后又再次下降。
用2,5-己二酮处理单组织MOC培养的神经球和肝脏物,并比较它们的类器官串联共培养表明,类器官串联共培养的组织比单培养的神经球和肝脏物的总和对2,5-己二酮更敏感(图7b)。在第9天和第10天用16m2,5 -己二酮处理的类器官串联共培养中,这种效应尤其明显。
此外,肝脏的单组织MOC培养比神经球对2,5-己二酮表现出更高的敏感性(图7c,d)。2,5-己二酮对MOC培养物也有剂量依赖性作用。
TUNEL/Ki67染色显示,在MOC类器官串联共培养的第0天和第14天,增殖细胞均匀分布在神经球和肝脏类器官中。然而,2,5-己二酮处理导致组织模型中凋亡细胞的明显增加(图8)。
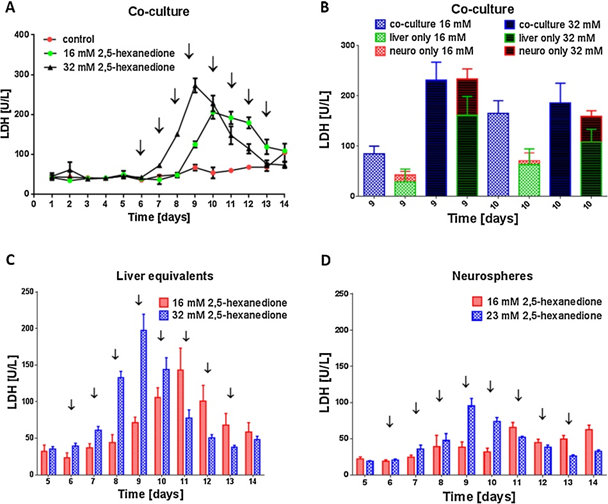
图7。未处理和处理过的培养基上清液中LDH的活性。分别用0、16 mM或32 mM 2,5-己二酮处理类器官串联共培养8天,从培养第6天开始。(A)类器官串联共培养培养基上清液中LDH活性的时间过程。(B)类器官串联共培养(类器官串联共培养)培养基上清液中LDH活性及培养第9天和第10天肝脏和神经元单组织MOC培养(仅肝脏,仅神经组织)中LDH活性之和。(C)肝脏单组织MOC培养和(D)神经球单组织MOC培养的培养基上清液中LDH活性的时间过程。(A)、(C)、(D):箭头表示物质应用;数据为四个独立培养的均值±标准差。
研究总结
微流体平台已被证明是一种稳定和强大的工具,以培养各种来源的组织在稳定下细胞外条件(Griffith et al., 1997;Huh,2012年;2012年;Materne et al., 2013)。本研究中使用的多器官芯片平台,在14天的动态流动条件下,在连接的介质电路中,可实现定义明确的神经球和肝脏的类器官串联共培养。细胞是活的,显示了组织特异性的标志物表达,并能够对2,5-己二酮的毒性损伤作出反应。据我们所知,这是第一次在微流体装置中进行超过14天的肝脏-神经球类器官串联共培养。
近年来,组织工程模式已经彻底改变了从二维体外测试系统向三维组织当量物质测试的转变。这些复杂的结构通常由多种细胞类型和集成技术(如生物材料科学)组成,是人类生理学的更接近模型,因此,导致更高的可预测性的临床结果(Wu,2010)。尽管这些系统在生产过程中往往更加耗时和成本密集,但它们能够在近生理细胞环境中对物质的作用模式进行高含量的分析(Esch et al., 2011;,2009年)。多种细胞类型整合成类器官结构,允许细胞相互作用并向成熟表型分化(Abu-Absi et al., 2004;Krause,2009)。我们之前开发了技术,以聚集分化的HepaRG细胞与HHSteC悬滴板。这些球状体被证明表达肝脏典型标记物,其水平与分化的单层细胞相当,甚至高于分化的单层细胞,即使球状体不再用DMSO刺激。细胞色素P450酶在球体的诱导表明一个代谢能力表现型。NT2细胞在搅拌悬浮培养系统中的聚集和神经元分化是由Serra(2009)开发的。在三周的分化过程中,这些神经球获得了成熟的神经元表型特征,如MAP2和泡状突触素染色,从而能够筛选潜在的神经毒性物质的作用。
将这两种复杂的组织结合在一个共同的介质循环中,以增加模型的复杂性,向预测不良结局途径的最终目标又迈进了一步。在标准的体外培养条件下,由于细胞的氧气和营养供应有限,三维组织模型往往会随着时间的推移而脱分化(Goral,2010; et al ., 2011)。这导致在临床前毒性试验中,物质对各自器官的影响的可预测性很低。然而,微流控系统中通过灌注的物质持续混合,可以更好地控制参数,如清除细胞分泌物和到达细胞的物理刺激。两种不同的器官系统结合在一个介质回路中,使组织通过分泌因子进行交流。此外,物质被肝脏代谢和随后到达神经系统的影响可以被研究。
未来系统物质检测最重要的要求之一是代谢产物到达次级器官系统的能力(Viravaidya和Shuler, 2004)。大多数微流体系统的流体与组织的高比率可能对有效的细胞通信构成挑战,因为大多数分泌因子在高中等容量中被稀释(Wikswo,2013年)。本研究中使用的MOC平台包括一个芯片上的微型泵,不需要外部介质储层。这使得芯片能够以非常低的中等容量运行,允许细胞相互作用。然而,神经球-肝脏类器官串联共培养的代谢活动(葡萄糖消耗和乳酸生成)表明,培养6天后达到了稳定的稳态。此外,在MOC中类器官串联共培养14天后,两种组织中仍发现有增殖细胞。
这些结果表明,小的中等体积能够维持两种组织,为细胞提供足够的营养和氧气。有趣的是,培养基中LDH活性在整个培养时间内保持不变,表明组织周转稳定。qRT-PCR结果显示,类器官串联共培养并没有改变组织的分化状态。
暴露于2,5-己二酮的类器官串联共培养的主要发现是,不仅可以区分两种不同剂量的毒性谱,而且与单一组织培养相比,类器官串联共培养对物质更敏感。我们假设,一个组织培养的坏死信号可能导致了第二种培养反应的增强。这一假设得到了这样一个事实的支持,即类器官串联共培养的敏感性增加主要是在暴露于较低浓度物质的培养中发现的。而暴露于较高浓度下的类器官串联共培养则没有这种强化效应。
总之,我们已经证明了高度分化的神经球和肝脏类器官可以在一个联合的培养基回路中类器官串联共培养超过14天,保持细胞分化的表型。此外,如上所述,我们已经表明,与单类器官培养相比,类器官串联共培养对2,5-己二酮的敏感性增加,这可能是由于组织-组织之间的通信。然而,还需要进一步的研究来阐明这一现象背后的机制。
北京佰司特贸易有限责任公司 (https://www.best-sciences.com):
类器官培养仪-HUMIMIC;质量光度计--OneMP;灌流式细胞代谢分析仪-IMOLA;便携式4通道SPR仪-P4SPR;蓝光/绿光LED凝胶成像;Nanocellect细胞分选仪-WOLF;微纳加工点印仪-NLP2000/DPN5000;
咨询
- 371
- 点赞
- 复制链接
- 举报









